
A overview of my scientific interests, the current research projects in which I am working on, and some other diversions.
Phenotypic plasticity – genetic bases and ecological effects

Project MORE: Phenotypic plasticity as a evolutionary driver in Moricandia
Project PREGEPES

Ecological and genetic processes producing speciation: the interplay between hybridization, poliploidy, and local adaptation in generalist plants.
Our starting hypothesis is that phenotypic changes associated with the interaction with pollinators can be involved in speciation even in species with a generalist pollination syndrome. A shift from yellow (typical in genus Erysimum) to purple petals might entail a shift in the relative abundance of the different types of pollinators, and might increase the visit rate of bees, which are highly efficient pollinators. However, these changes do not suffice to ensure reproductive isolation. Hence, other factors such as local adaptation, especially in the earliest ontogenetic stages might have played a relevant role in the radiation and diversification of the genus. Lastly, hybridization and polyploidy would foster and accentuate the differentiation process, inducing speciation. The working hypothesis is that the high rates of speciation of Erysimum are the joint result of all these processes.
Project STRUGEN
Structured Generalization: evolutionary dynamics at fine spatial scales in a generalist system.
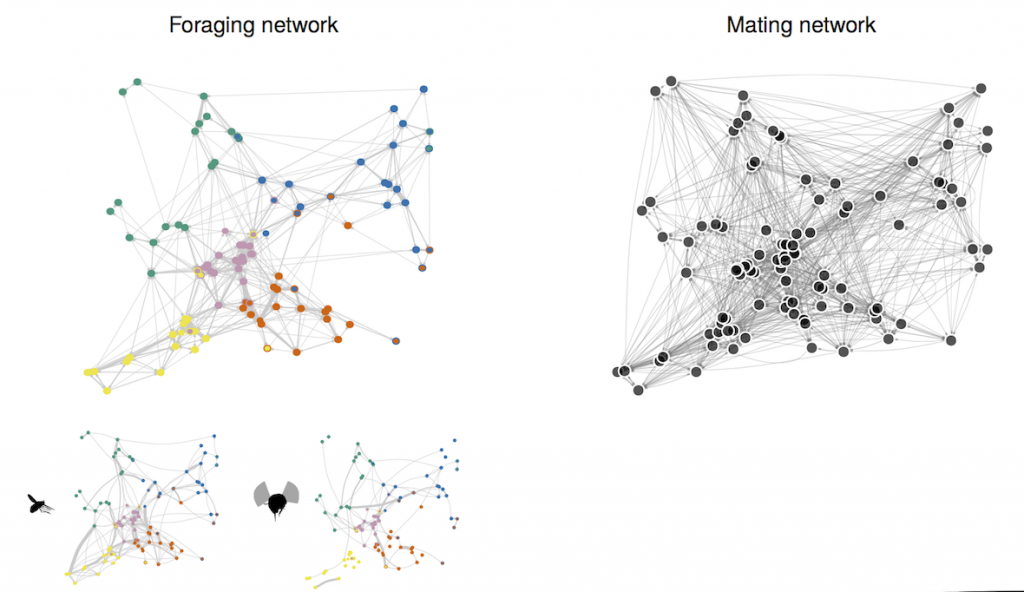
In pollination generalist systems, all plant genotypes in a population are assumed to interact with random subsets of the overall pollinator pool. In this project we explored the idea that the generalization degree of a given plant could be non random, but related with some intrinsic and extrinsic factors, such as its microenvironment, its spatial location in the population, and most important, its phenotype and genotype. Plants exhibiting different values for those factors would attract different subset of pollinators, and plants showing different values of those traits would share similar subgroups of pollinators.
These inter-individual differences could promote a structured pattern of interaction among the plant and its pollinators, a phenomenon that we call structured generalization. We propose that structured generalization occurs when there are non-random inter-individual differences in generalization and the intraspecific pollination and mating networks at population level are divided in subgroups of plants sharing similar pollinators (i.e., network is clustered).
My scientific interests are mainly developed through the collaboration with two research groups:
Evoflor is an informal team of scientists with the common interest in explore the evolutionary biology of pollination interactions.
We use an wide range of approaches, including observational and experimental field data, molecular marker analysis, sequence data, greenhouse experiments, and in silico modeling to analyze the diffuse co-evolution between flowers and their pollinators. Our interdisciplinary research is centered in the role of the corolla shape as signal to pollinators. We are using some Brassicaceae species, mainly from the Erysimum genus, as our focal and model species.

The B-chromosomes, also known as accessory or supernumerary chromosomes, are dispensable chromosomes found in a wide variety of species, from fungus to mammalian animals. These chromosomes usually reduce the fitness of their carriers, thus behaving as genomic parasites. B-chromosomes usually persist in natural populations thank to mechanisms that improve their transmission, over-surpassing the Mendelian ratios.

At the Evolutionary Genetic Group at the University of Granada, we are researching the complete life cycle of these parasites, with a special focus on the B-chromosome system of the grasshopper Eyprepocnemis plorans. This grasshopper harbors an extremely widespread polymorphic B-chromosome system, with different B-chromosomes found in almost all circum-Mediterranean and Caucasian populations hitherto sampled.

B chromosomes in this species have been shown to evolve through several stages of parasitic and near-neutral nature, presumably because of an arms race between the standard (A) and B chromosomes. This intragenomic conflict can either be solved with the extinction of the neutralized B chromosome or, more interestingly, with the replacement of this neutralized B-chromosome by a new mutant version, being parasitic again and thus prolonging B chromosome life.
My research is supported through grants from several institutions
