
Etimológicamente, la palabra Halloween proviene de la contracción de all hallow’s eve, es decir, la víspera del día de todos los santos. Por simplificar, esta festividad que se celebra los primeros días de cada mes de noviembre, tiene origen celta, pero adquirió un sentido religioso a través del sincretismo cristiano y cada grupo la adaptó a su realidad cultural (Día de Muertos, Día de Todos los Santos, Día de los Fieles Difuntos). Hasta que llegó la globalización… En efecto, estos días, la calle y los comercios son un continuo recordatorio de cómo esta tradición del folclore popular americano se sigue haciendo un hueco en nuestra sociedad: los disfraces, el truco o trato y, por supuesto, las calabazas huecas con formas fantasmagóricas. De estas últimas quería hablar hoy.
 Calabazas preparadas para Halloween.
Calabazas preparadas para Halloween.
Es posible que, como consecuencia del empuje de esta festividad, dejemos pasar por alto que no todas las calabazas son de color naranja intenso. Basta dar un paseo por cualquier frutería para encontrarlas naranja pálido-crema (la mayoría), o verde (cada vez menos frecuentes, al menos en grandes superficies).
 Calabazas verdes y naranja pálido en la estantería de mi frutería habitual un día cualquiera.
Calabazas verdes y naranja pálido en la estantería de mi frutería habitual un día cualquiera.
Los compuestos responsables de esta coloración son los carotenoides, terpenos que dan color amarillo, anaranjado o rojo a muchas especies vegetales. A continuación, se muestra un esquema de la ruta metabólica de síntesis de dichos pigmentos (en concreto, la calabaza es rica en β–caroteno, fuente muy importante de vitamina A en nuestra dieta):
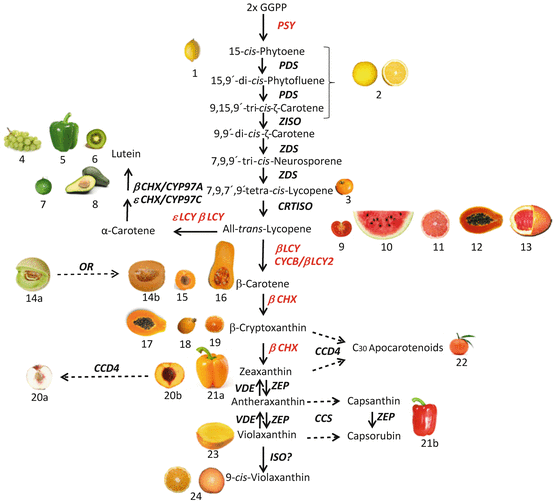 Fuente: Lado et al., 2016
Fuente: Lado et al., 2016
Como se deduce de este esquema, se trata de una ruta metabólica compleja en la que participan numerosos genes. De hecho, es habitual que varios genes formen parte de un mismo proceso bioquímico, contribuyendo así a la expresión de un rasgo concreto. En dicho caso, se considera que hay interacción entre genes. Esta interacción puede llevarse a cabo de distintas maneras; una de ellas, la epistática, tiene lugar cuando uno de los genes enmascara el efecto del otro. En Genética, la ruta metabólica responsable del color final de la calabaza es ejemplo clásico de interacción génica epistática. Veámoslo en detalle:
En concreto, la coloración depende, entre otros (*), del gen Mature Orange, que puede expresar un fenotipo naranja (Co_) o verde (coco). El gen Inhibitor of Mature Fruit Color, tiene un alelo dominante (Ic) que actúa como inhibidor del gen anterior y se podría considerar que ejerce un efecto epistático sobre él, ya que impide los alelos Co/co expresen sus correspondientes fenotipos tal y como muestra el siguiente esquema:
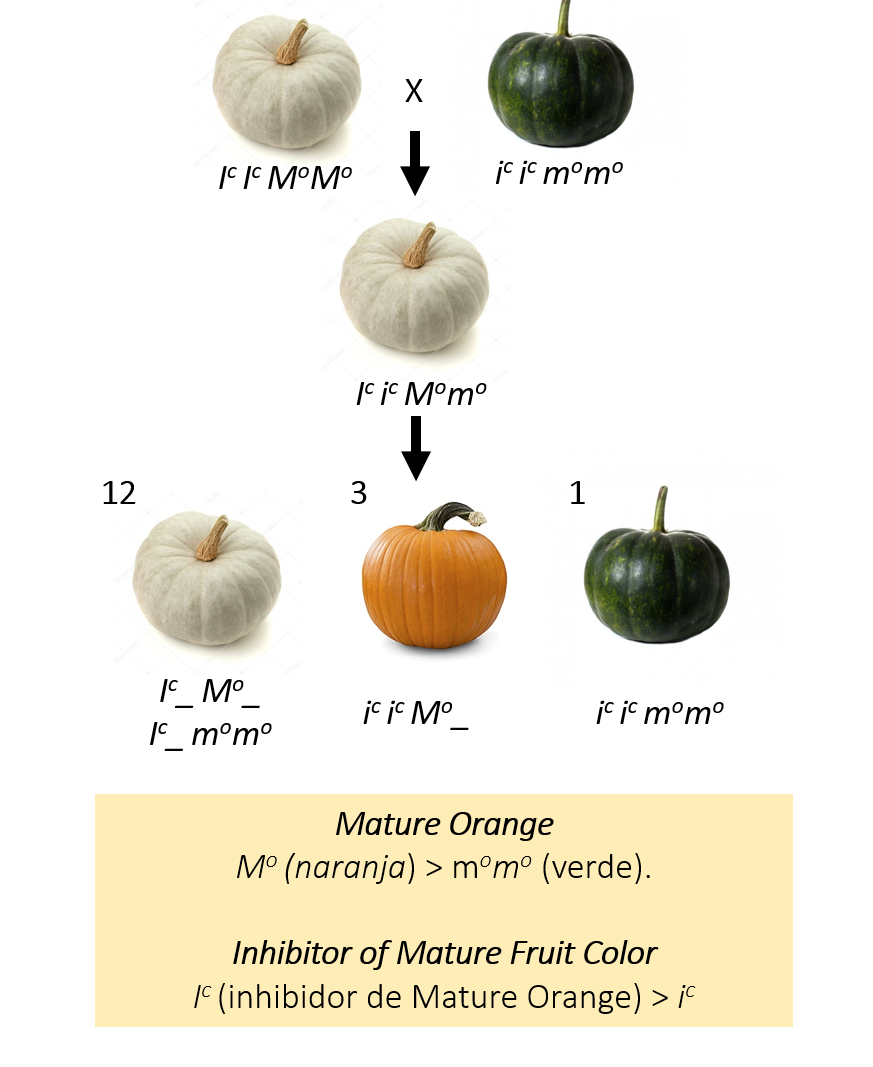
Ésta se considera una epistasis simple dominante. Simple, porque el gen epistático (Inhibitor of Mature Fruit Color) ejerce su efecto unidireccionalmente sobre el gen hipostático (Mature Orange) y nunca al revés. Dominante, porque el efecto epistático se produce cuando aparece el alelo dominante del gen epistático (en este caso, Ic). Como puede observarse, esta interacción produce una alteración de las proporciones 9:3:3:1 (esperables de un cruzamiendo entre dos dihíbridos) hacia un 12:3:1, ya que tanto los individuos Ic_Mo_ como los Ic_momo tienen una forma del gen inhibidor activa y, por lo tanto, expresan el mismo fenotipo (ausencia de pigmento). Esto explicaría por qué en el mercado predominan las calabazas con colores anaranjados sobre las de color verde.
(*) A pesar de tratarse de un ejemplo habitual en los libros de texto, no he podido encontrar el nombre concreto de los genes implicados en esta epistasis. Es por ello, que consultando la lista de genes descritos para esta especie, he seleccionado una pareja génica responsable de la coloración y a la que podría atribuirse una relación epistática.
Bibliografía:
- Harry S. Paris H.S. (2005). The genes of Pumpkin and Squash. HortScience, 40(6):1620–1630.
- Lado J., Zacarías L., Rodrigo M.J. (2016) Regulation of carotenoid biosynthesis during fruit development. In: Stange C. (eds) Carotenoids in Nature. Subcellular Biochemistry, vol 79. Springer, Cham.
- Sun H., Wu S., Zhang G., Jiao Ch., Guo S. et al. (2017). Karyotype stability and unbiased fractionation in the paleo-allotetraploid Cucurbita genomes. Molecular Plant, 10(10):1293-1306.